
This week’s BEACON Researchers at Work blog post is by MSU postdoc Jeff Morris.
 As the 18th century dawned over Europe, pretty much everybody believed the world was as it was because of a mysterious divine plan. But during the period known as the Enlightenment, thinkers began working out the mechanics that structured both nature and human society. By century’s end many of the fundamental processes of physics and chemistry had been elucidated. But the greatest leap ahead in the history of biology – the discovery by Charles Darwin of evolution by natural selection – wouldn’t happen until the middle of the 19th century, and it would owe as much to the study of economics as to anything in the preceeding century’s life sciences.
As the 18th century dawned over Europe, pretty much everybody believed the world was as it was because of a mysterious divine plan. But during the period known as the Enlightenment, thinkers began working out the mechanics that structured both nature and human society. By century’s end many of the fundamental processes of physics and chemistry had been elucidated. But the greatest leap ahead in the history of biology – the discovery by Charles Darwin of evolution by natural selection – wouldn’t happen until the middle of the 19th century, and it would owe as much to the study of economics as to anything in the preceeding century’s life sciences.
Adam Smith, an 18th century Scottish philosopher, was struck by how economics obeyed natural-like laws despite the often capricious and irrational behavior of economic actors. Smith envisioned an intrinsic interrelation between producers and consumers that pulled economic behavior toward certain norms as if “led by an invisible hand.” The net result of many financial actors behaving selfishly is a well-regulated, self-organized system: the parts don’t have any intention of working together, but screw up and do it anyway.
Young Charles Darwin was influenced by many, many Enlightenment thinkers, but it’s hard not to notice how similar the self-organization of nature by natural selection is to Smith’s ideas about economics. This Spectator article from 2009 covers Darwin’s economic influences in depth. It also uses one of my favorite weird words, “defenestrate”:
“Ideas evolve by descent with modification, just as bodies do, and Darwin at least partly got this idea from economists, who got it from empirical philosophers. Locke and Newton begat Hume and Voltaire who begat Hutcheson and Smith who begat Malthus and Ricardo who begat Darwin and Wallace. Before Darwin, the supreme example of an undesigned system was Adam Smith’s economy, spontaneously self-ordered through the actions of individuals, rather than ordained by a monarch or a parliament. Where Darwin defenestrated God, Smith had defenestrated government.” – Matt Ridley
Marketplace analogies are common in biology: here are two dealing with microbial evolution. And when applied to human social organization, evolution often draws the same criticisms as free market policies: can “nature red in tooth and claw” produce cooperation and charity, or must we rely on a benevolent dictator to give us those happy institutions?
The chief problem with evolving cooperation is the tragedy of the commons. Briefly, if cooperation has a cost, then a non-cooperator that can still get the benefits of cooperation will always have a fitness advantage over cooperators. Theoretically, this advantage will always exist even if the breakdown of cooperation totally trashes the environment. We know the tragedy doesn’t always happen because we see organisms in nature working together – but how does evolution escape it?
Bill Hamilton proved that self-sacrifice could evolve by natural selection if the recipients of the sacrifice were related to the sacrificer – something we’ve come to call kin selection. In the microbial world, kin selection can happen when microbes live in close physical association with each other. Since microbes reproduce by simple division, as long as they don’t move around they’re likely to be surrounded by close relatives. Therefore, if a cell spends resources to make a product that leaks into the environment, the cells most likely to benefit are also close relatives.
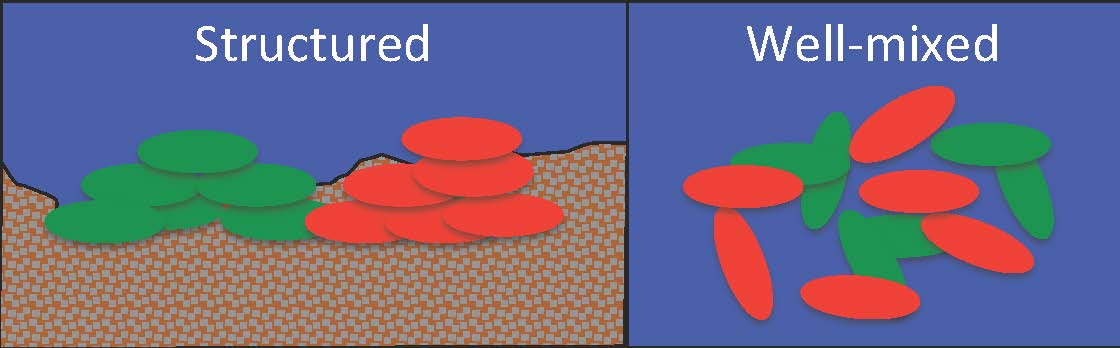
Bacteria living in a spatially structured environment like a seabed (left) are more likely to be related to their neighbors than the same organisms living in open, well-mixed water (right). Classic kin selection can happen in the scenario on the left, but not in the one on the right.
But kin selection can’t explain all the cooperation we observe in the microbial world. In fact, we see tons of it in the open ocean, whose turbulent waters are about as close as you’re likely to get to no spatial structure at all. The oceans are full of cells that are dependent on unrelated cells to make crucial metabolites, or in some cases to clean up environmental toxins. In order to understand how this arrangement evolved, we need to consider two theories:
- Streamlining theory arose from work with the omnipresent marine bacterium SAR11 done mostly by Steven Giovannoni’s group at Oregon State. It maintains that there is intense selection pressure on microbes to reduce the sizes of their genomes when nutrients are limiting and populations are large. During this process of gene loss, many cells lose the ability to perform vital functions and become dependent on neighboring cells that have retained those functions.
- The Black Queen Hypothesis, or BQH, which I originally proposed in my PhD thesis and then worked out more rigorously in a 2012 mBio paper with Erik Zinser and Richard Lenski, proposes that streamlined cells can get away with losing important functions as long as those functions leak their products into the environment. Like players of the card game Hearts want to get rid of the black Queen of Spades because of her high point cost, streamlining cells want to get rid of leaky functions. However, some cells have to end up holding the Black Queen, because once the leaked products are rare enough, additional streamlining moochers won’t have an advantage. BQH evolution thus produces communities of function-performers, or helpers, and moochers, or beneficiaries. These communities have the appearance of cooperation/altruism, but they arise by normal selfish Darwinian natural selection.
We originally proposed the BQH to explain how the marine photosynthetic bacterium Prochlorococcus had become dependent on its neighbors for protection from hydrogen peroxide, a toxin that is constantly produced in natural waters exposed to sunlight. Because peroxides move freely across cellular membranes, any cell protecting itself by breaking down the peroxides also lowers the environmental concentration of peroxide, unavoidably helping any mooching neighbors. This leakiness makes peroxide detoxification a Black Queen function and explains how Prochlorococcus can get away with not protecting itself.
In order to test the predictions of the BQH, we used a wimpy Prochlorococcus-like E. coli mutant that had all of its anti-peroxide defenses knocked out. We then gave this mutant a plasmid – an accessory piece of DNA – that allows the cells to make the peroxide-destroying enzyme KatG and evolved the resulting strain under strong, peroxide-generating light for 1,200 generations, or about 150 daily transfers of the cultures into fresh batches of growth medium.
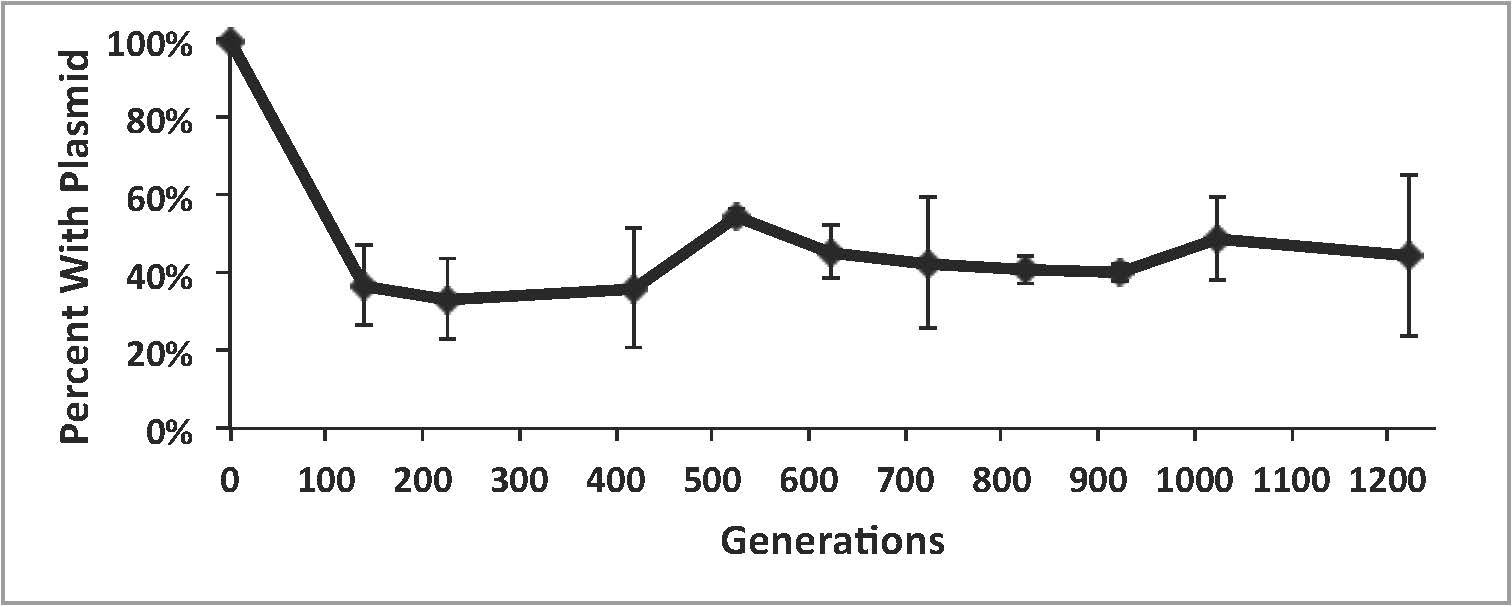
Percentage of the evolving E. coli populations retaining the KatG plasmid. Values are means of 3 replicately evolved populations; error bars are standard deviations.
Even though plasmid-free cells could barely survive on their own under the lights, they nevertheless evolved and stably co-existed with their plasmid-containing ancestors throughout the 1,200 generations (see above). Moreover, while the helpers underwent a number of evolutionary changes, there was no evidence that they were trying to be “stingy” with their production of KatG in order to outcompete the beneficiaries. Just the opposite – the evolved helpers made more KatG than their ancestors.
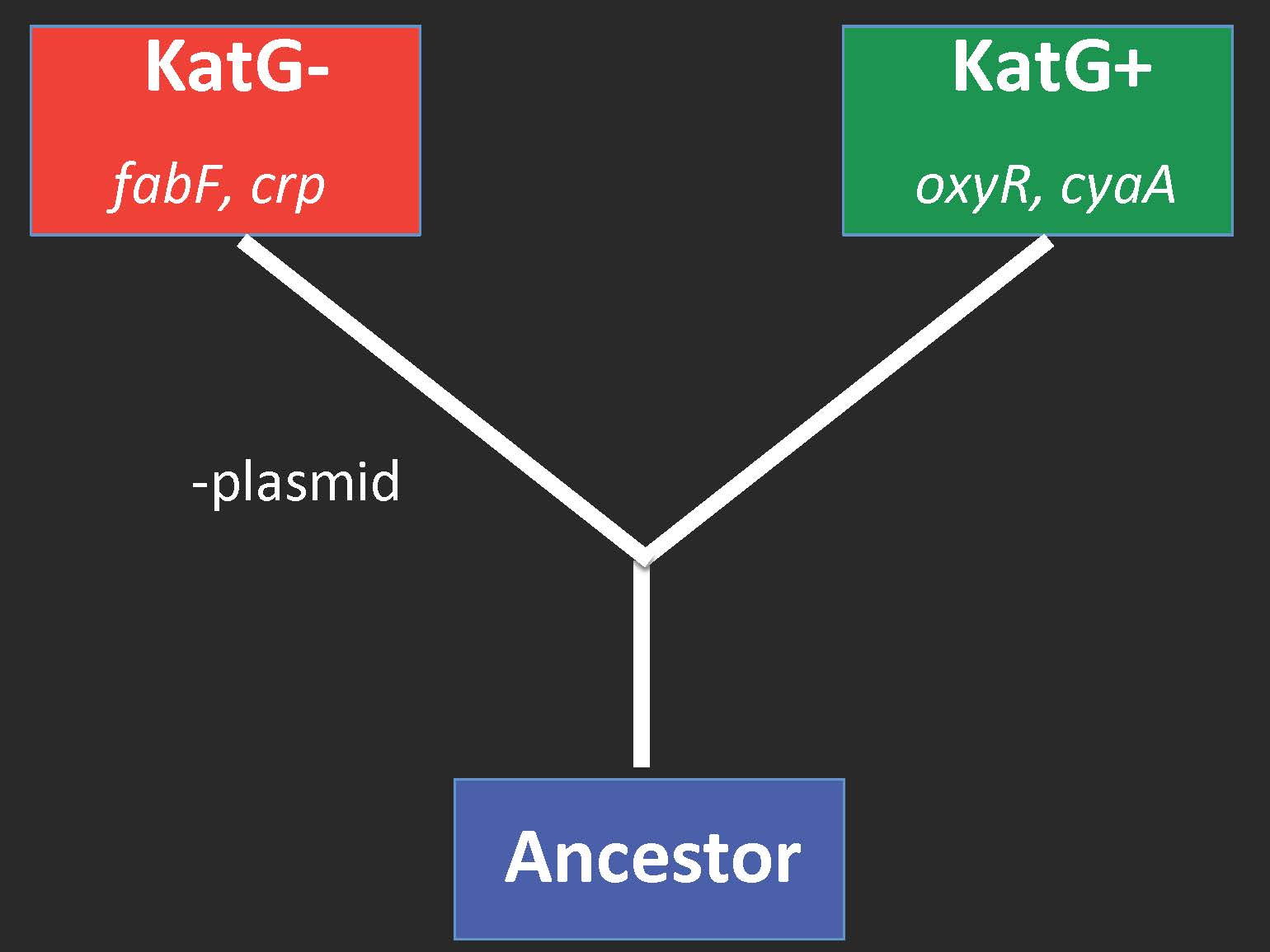
Family tree of genes that mutated in more than one of the three evolved populations.
Not only did the two forms coexist, there’s also evidence that they diverged into different “species.” When we sequenced the genomes of helper and beneficiary clones taken from three replicate evolved lines, we found two mutated genes common to all helper lines, and a completely different set of two mutated genes in all beneficiary lines (see above). This indicates that the early choice to become a beneficiary fundamentally changed the adaptive landscape of these cells, meaning different mutations are adaptive for helpers than for beneficiaries. This is a barrier to gene flow, and based on the ecological species concept, these two types represent different species (or more properly for bacteria, ecotypes) – evolved in 150 days under a simple BQH regime!
Starting with a single, clonal population, Black Queen evolution produced an ecosystem with 2 distinct types, one of which was apparently altruistically helping the other. Of course, we know that they aren’t helping because they care about the beneficiaries; they just can’t avoid doing so. Each player is maximizing its own selfish advantage at the other’s expense, but the nature of the leaky function prevents either from winning the game and taking sole control of the evolutionary medium.
The BQH thus acts as an invisible hand stablizing the ecosystem and forcing the two types to play nice with each other. Are there any important differences between this kind of inadvertant cooperation and what we might think of as “intentional” cooperation? Maybe, maybe not. Either way, though, it’s clear that the BQH equilibrium can keep varieties of closely related organisms co-existing for a long time, and, unable to get rid of their pesky partner, it’s possible that other forms of cooperation might evolve in time.
Human attempts to manipulate complex systems, be they economies or ecosystems, are often studies in disaster. The invisible hand of the Black Queen shows us one of many ways that blind nature is often a better engineer than any intelligent designer.
References:
Jeff Morris is a NASA Astrobiology Institute postdoctoral fellow working in Richard Lenski’s lab at Michigan State. He will be starting as an assistant professor in the Biology Department at the University of Alabama in Birmingham in January. Jeff (@ASDarwinist) blogs about science, politics, and heavy metal at antisocialdarwinist.com.






























{kind=link}