

MSU graduate student Andrea Morrow
This post is by MSU graduate student Andrea Morrow.
People often associate certain physical adaptations with an animal’s temporal niche, i.e. daily activity pattern. For example, some nocturnal animals have larger eyes, larger corneas, or higher concentrations of rod cells than diurnal animals [1]. These traits are valuable for animals that are awake and active at night because they augment the collection and utilization of what little light is available, which is advantageous when it comes to avoiding predators and finding resources. While these adaptations may be valuable, nervous tissue involved in processing sensory information requires a large amount of energy to develop, maintain, and power [2]. For this reason, it might not be a great idea for a nocturnal animal to invest extensive amounts of energy into processing complex visual information, when the amount of information obtained will always be restricted due to lack of light. Let’s think of the flip side to that. Would a diurnal animal, with plenty of light available, profit more than a nocturnal animal from expending substantial energy in visual processing?
In general, if the energetic cost is high, then the reward should also be high, or else the trait will not be maintained [3]. We can apply this concept of costs and benefits to all sensory processing. Brain size is constrained by energetic costs [4], and therefore we expect energetic trade-offs when it comes to investments in specific areas or tasks. How do nocturnal vs diurnal animals utilize visual, tactile, olfactory, and auditory cues from their environment? Are some more heavily relied upon by diurnal animals, and less by nocturnal animals? If, as I suspect, diurnal animals invest more in visual processing than nocturnal animals do, does it come at a cost in the form of other sensory processing?
Dr. Barbara Lundrigan (Department of Integrative Biology), Dr. Laura Smale (Department of Psychology), and I are trying to answer some of these questions by examining multiple sensory processing structures within the brain. We want to determine whether investment trade-offs between olfactory, visual, and auditory processing occur following evolutionary shifts in activity pattern.
Focal group
Determining the effect of a single factor in an observational study can be a heady task, especially when it comes to complex brain structures. Ecological, behavioral, and phylogenetic factors all work in shaping brain morphology and functionality. We are sampling widely across the order of Rodentia (which includes more than 2,000 extant species) in order to include species that vary in life history, behavior, environment, and evolutionary history. Within this broad framework, we are sampling closely related species within multiple subfamilies where temporal niche transitions have occurred independently. For example, with our colleague Dr. Paul Meek, we are sampling Australian bush rats (nocturnal) and Australian swamp rats (diurnal) from the family Muridae. We are also sampling Southern flying squirrels (nocturnal) and North American red squirrels (diurnal) from the family Sciuridae. Both of these groups have independently experienced activity pattern shifts. We want to capture as many independent evolutionary shifts as possible, as well as including animals that exhibit activity patterns other than strict nocturnality and diurnality. Our working list currently includes about 30 different species from 10 subfamilies, representing several independent shifts. We have collected tissue samples from 13 species and over 80 individuals to-date.
Structures of interest
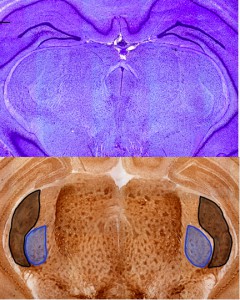
Two coronal sections of an Australian rat. The top image is stained with Cresyl Violet. The bottom image is a section stained with Acetylcholinesterase. The Medial Geniculate Nucleus (MGN) is outlined and shaded in blue and the Lateral Geniculate Nucleus (LGN) is outlined and shaded in black.
The goal is to quantify the investment in olfactory, visual, and auditory processing. For olfactory processing, we are measuring the weight of the olfactory bulbs, relative to the weight of the entire brain, in each individual. For visual and auditory processing, we are measuring the volume of the Lateral Geniculate Nucleus and specific layers of the Superior Colliculus, which are visual processing structures, and the Medial Geniculate Nucleus and the Inferior Colliculus, which function in auditory processing. We do this by cryosectioning the frozen, unfixed brain tissue of each animal. The tissue is stained using two types of stain (Cresyl Violet and Acetylcholinesterase), microphotographs of each tissue section are taken and uploaded to the computer, and then each structure is delineated. These histological methods allow us to measure the area of each structure on each section of brain tissue, which we then use to calculate volume.
Our objective is to better understand how animals utilize the different sensory cues in their environment, as well as how investment in the processing of that information changes with the evolution of different activity patterns. While we don’t have data on the visual and auditory structures yet, we have collected some data on the olfactory bulbs. While the data are preliminary and represent only 13 species, with 1-6 individuals each, differences in relative olfactory bulb size between nocturnal and diurnal species are already apparent. The nocturnal species possess larger olfactory bulbs, relative to brain size, than the diurnal species. In addition to adding more species, we plan to look more thoroughly at the olfactory system in relation to activity pattern in the future.
I’ve always been interested in how organisms adapt to their environments and how their morphology and physiology are shaped (or constrained) by their evolutionary history and ecology. It is intriguing to imagine one environment (spatially) being separated temporally simply by the rotation of the earth and the ensuing cycles of light. The temporal niche presents many unanswered questions. What changes occur when animals adapt to a new temporal niche? Are these changes consistent and predictable? What conditions permit or restrict temporal partitioning? Is competition a driving force of this type of niche shift? Must there be plasticity, or flexibility, in activity pattern present for such shifts to occur, and if so, at what level? I’m hoping that, in time, I will be able to answer at least a few of these questions.
References
[1] Kirk EC. 2006. Effects of activity pattern on eye size and orbital aperture size in primates. Journal of Human Evolution. 51:159-170.
[2] Mink JW, Blumenschine RJ, Adams DB. 1981. Ratio of central nervous system to body metabolism in vertebrates: its constancy and functional basis. American Physiological Society. 241:R203-R212.
[3] Niven JE, Laughlin SB. 2008. Energy limitations as a selective pressure on the evolution of sensory systems. Journal of Experimental Biology. 211:1792-1804.
[4] Barrickman NL, Lin MJ. 2010. Encephalization, expensive tissues, and energetics: An examination of the relative costs of brain size in Strepsirrhines. American Journal of Physical Anthropology. 143:579-590.
