
This week’s BEACON Researchers at Work post is by MSU graduate student Raffica La Rosa.
 Novel traits differ qualitatively from the characters from which they arise, and are generally thought to be adaptive. I study adaptive novel traits by combining studies of present-day natural selection to identify which traits are likely adaptive, and phylogenetic comparative analyses to understand the past evolution of those traits. My study system is the milkweed genus Asclepias. Milkweeds have amazing flowers that are distinct from all other other flowers on Earth. They have unique floral traits that likely influence how they interact with their insect pollinators, but little is known about how the floral traits might be adaptive.
Novel traits differ qualitatively from the characters from which they arise, and are generally thought to be adaptive. I study adaptive novel traits by combining studies of present-day natural selection to identify which traits are likely adaptive, and phylogenetic comparative analyses to understand the past evolution of those traits. My study system is the milkweed genus Asclepias. Milkweeds have amazing flowers that are distinct from all other other flowers on Earth. They have unique floral traits that likely influence how they interact with their insect pollinators, but little is known about how the floral traits might be adaptive.
Milkweed flowers have a number of unusual floral structures. Typically, an angiosperm flower is made up of four whorls. The sepals are the outermost whorl, are often green, and typically form the bud before the flower opens. The next whorl in is made up of petals, which we usually think of as the colorful, attractive part of the flower. The innermost whorls consist of the reproductive organs—the stamen produce pollen (male) and the carpels contain ovules (female). Milkweeds in the genus Asclepias have sepals and petals, but their male and female whorls have fused together and are almost unrecognizable.
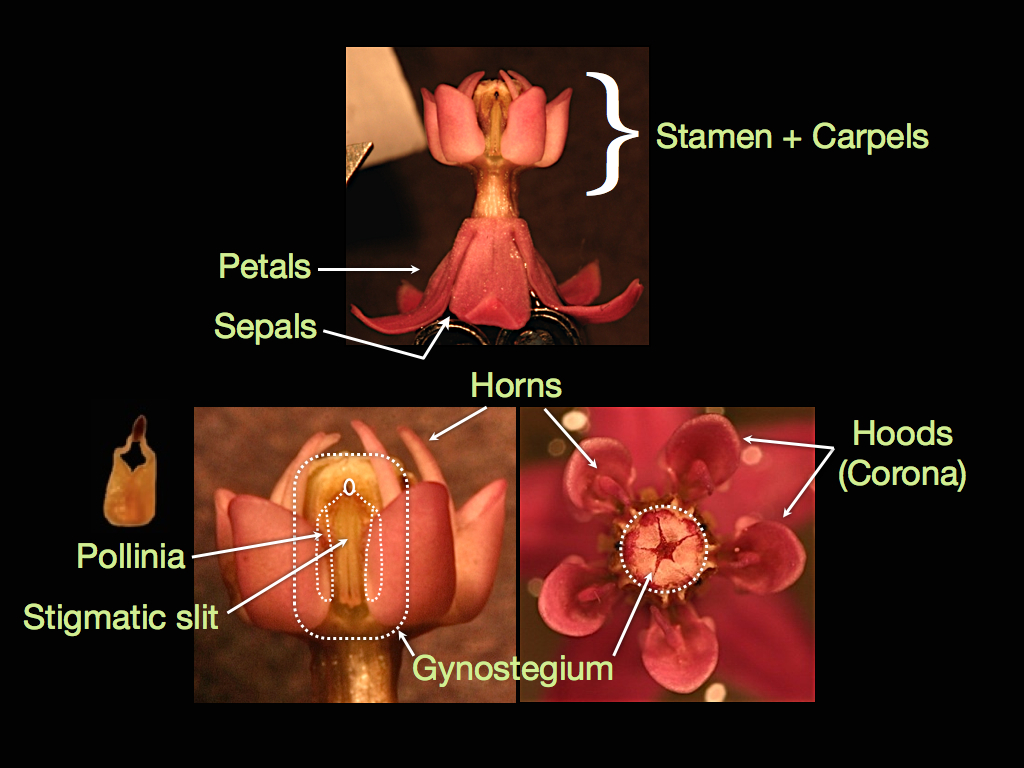 In the center of the flower is the gynostegium, which forms a chamber containing two carpels. Rather than having loose pollen that might stick to pollinators, the pollen has been clustered together into five pairs of waxy pollen sacks called pollinia. The pollinia reside in the walls of the gynostegium between nectar-holding hoods that often have horn-like protuberances. The exposed dark gland (corpusculum) that attaches adjacent pollen sacks has a tapered slit, like the back of a hammer, that catches onto the hairs, claws, or mouthparts of pollinators, and allows the pollinia to be slipped out and transferred between flowers.
In the center of the flower is the gynostegium, which forms a chamber containing two carpels. Rather than having loose pollen that might stick to pollinators, the pollen has been clustered together into five pairs of waxy pollen sacks called pollinia. The pollinia reside in the walls of the gynostegium between nectar-holding hoods that often have horn-like protuberances. The exposed dark gland (corpusculum) that attaches adjacent pollen sacks has a tapered slit, like the back of a hammer, that catches onto the hairs, claws, or mouthparts of pollinators, and allows the pollinia to be slipped out and transferred between flowers.
 For pollination to occur, a single pollinium must be deposited into one of five slits around the outside of the gynostegium that lead to the central chamber. Once there, each pollen grain can grow a pollen tube to fertilize an ovule. The pollinium contains enough pollen grains to fertilize all of the ovules within a carpel, so the milkweed fruit (pod) that develops often contains 40-200 seeds that all share the same father. This can be very convenient for researchers, such as myself, who want to study natural selection through female fitness and male fitness, which is rarely done in plants. Measuring male fitness in plants is often very difficult because loose pollen from many individuals is easily jumbled, resulting in a fruit containing seeds sired by many different fathers; in milkweeds however, I can collect a pod, sprout just one seed, run genetic tests to figure out paternity, and then know the paternity of all of the seeds in that pod. This gives me the ability to find the paternity of up to 100% of the seeds in a population by only sampling about 3% of them!
For pollination to occur, a single pollinium must be deposited into one of five slits around the outside of the gynostegium that lead to the central chamber. Once there, each pollen grain can grow a pollen tube to fertilize an ovule. The pollinium contains enough pollen grains to fertilize all of the ovules within a carpel, so the milkweed fruit (pod) that develops often contains 40-200 seeds that all share the same father. This can be very convenient for researchers, such as myself, who want to study natural selection through female fitness and male fitness, which is rarely done in plants. Measuring male fitness in plants is often very difficult because loose pollen from many individuals is easily jumbled, resulting in a fruit containing seeds sired by many different fathers; in milkweeds however, I can collect a pod, sprout just one seed, run genetic tests to figure out paternity, and then know the paternity of all of the seeds in that pod. This gives me the ability to find the paternity of up to 100% of the seeds in a population by only sampling about 3% of them!
With this handy feature of milkweed flowers, I can measure selection on floral traits through female fitness and male fitness separately to see if the unusual floral traits of milkweeds function more to help the plants produce more seeds (female fitness), or sire more seeds (male fitness). To do so, I just need just three things to run a selection gradient analysis: trait measurements, female fitness measurements, and male fitness measurements.
 To collect trait measurements, I first determine which floral traits might be influencing the attraction, reward, and efficiency of pollinators, since Asclepias species depend on insect pollinators to transfer their pollen. After observing the flowers and their interactions with pollinators in nature, I choose floral traits that I think might be influencing pollinators, and ultimately affecting fitness. For instance, the size of the hoods and gynostegium most certainly affect the visibility of the flowers, the dimensions of the hood could affect the volume of the nectar reward, and the horns and spacing between hoods could influence how easily pollinators remove and deposit pollinia. To measure the traits, I collect several flowers per plant in the population, digitally photographed them, and later measure them from the photographs.
To collect trait measurements, I first determine which floral traits might be influencing the attraction, reward, and efficiency of pollinators, since Asclepias species depend on insect pollinators to transfer their pollen. After observing the flowers and their interactions with pollinators in nature, I choose floral traits that I think might be influencing pollinators, and ultimately affecting fitness. For instance, the size of the hoods and gynostegium most certainly affect the visibility of the flowers, the dimensions of the hood could affect the volume of the nectar reward, and the horns and spacing between hoods could influence how easily pollinators remove and deposit pollinia. To measure the traits, I collect several flowers per plant in the population, digitally photographed them, and later measure them from the photographs.
To measure fitness, I collect all of the pods in the population and record how many each plant produces. Later, I count the number of seeds in each pod. From these data alone, I can quantify female fitness, because I can say how many seeds each plant has produced. Measuring male fitness is a much longer process that starts by sprouting one seed from each of the pods. Once the seedlings are large enough, I collect them and extract their DNA. During the summer, I also collect leaf tissue from every possible parental plant in the population and extract DNA, so that I can match the offspring to their parents. I already know who the mothers are, but I can use genetic paternity tests to identify the fathers.
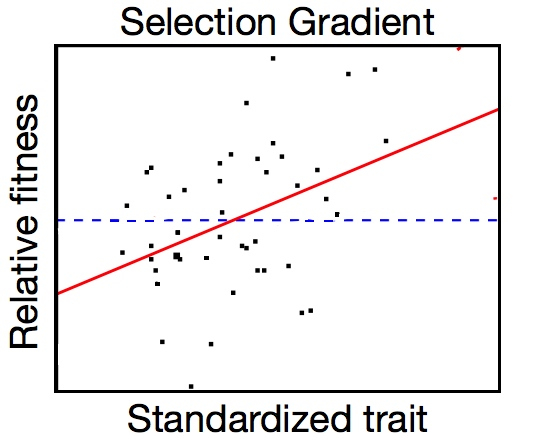
Regression of relative fitness onto a trait; the slope of the fitted line is the selection gradient. Each point represents an individual within the population.
I measure natural selection on each of the traits by using a multiple regression to regress relative fitness onto all six of the traits at once to account for any correlations between traits. The resulting coefficients are the selection gradients. Positive selection gradients mean that individuals with larger trait values will have higher fitness, and negative selection gradients mean that plants with smaller values of that trait have higher fitness. The larger the absolute value of the selection gradient is, the stronger selection is. Finding selection on a trait is a large first step toward knowing if a trait is adaptive.
 For more information about Raffica’s work, you can contact her at larosara at msu dot edu.
For more information about Raffica’s work, you can contact her at larosara at msu dot edu.
