
This week’s BEACON Researchers at Work blog post is by Michigan State University postdoc Sarah Bodbyl.
 “Bees do have a smell, you know, and if they don’t they should, for their feet are dusted with spices from a million flowers.” ― Ray Bradbury, Dandelion Wine
“Bees do have a smell, you know, and if they don’t they should, for their feet are dusted with spices from a million flowers.” ― Ray Bradbury, Dandelion Wine
Recent and dramatic declines of pollinating insects have been observed all over the world by scientists and amateurs alike. Human-induced environmental changes, such as habitat loss, climate change, invasive species introductions, pesticide use, and novel disease transmission are all thought to be contributors to this global pollinator decline. Since many (at least 180,000) plant species rely on pollinators to reproduce via seeds and fruit, pollinator declines have folks worried about both wild and agricultural plant populations.
One of my research areas investigates the question, what happens to plants when they lose their pollinators? Specifically, I want to know if plants experiencing rapid loss of their pollinators are able to adapt and maintain viable populations by increasing their ability to reproduce without pollinators.
Mating system evolution
Flowering plants, or angiosperms, have evolved numerous reproductive strategies to overcome their sessile nature. Outcross fertilization (outcrossing) is a sexual reproduction strategy occurring when pollen from the flower of one plant is transferred to the flower of another plant. Self-fertilization (selfing, or autogamy) occurs when pollen from one flower fertilizes the ovaries of the same flower, or another flower within the same plant. Some plants are also capable of asexual reproduction via apomixis or vegetative cloning, but my research focuses on sexual strategies. Mating system is the degree of allocation by a plant population or species to each reproductive strategy. Some plants exclusively outcross, some exclusively self, and others employ both in varying degree; termed mixed mating. Both outcrossing and selfing can occur either with or without the aid of pollinators, but many flowering plants have evolved mutualistic relationships with pollinators to facilitate outcrossing.
 As in most variable traits found in nature, plant mating systems are not fixed and evolve in response to selective pressures. The evolutionary transition from outcrossing to selfing has occurred repeatedly in angiosperms and is associated with changes in floral biology, life history, and ecology. Some evolutionary biologists have predicted that plant populations that lose their pollinators may be capable of evolving increased selfing capacity, a mechanism of reproductive assurance. However, this transition has never been directly observed taking place in a plant population in response to pollinator loss, so I decided to try to recreate the process in an experimental setting.
As in most variable traits found in nature, plant mating systems are not fixed and evolve in response to selective pressures. The evolutionary transition from outcrossing to selfing has occurred repeatedly in angiosperms and is associated with changes in floral biology, life history, and ecology. Some evolutionary biologists have predicted that plant populations that lose their pollinators may be capable of evolving increased selfing capacity, a mechanism of reproductive assurance. However, this transition has never been directly observed taking place in a plant population in response to pollinator loss, so I decided to try to recreate the process in an experimental setting.
Experimental evolution of mating system in monkeyflowers
To determine the immediate effects of pollinator loss on a plant population, my doctoral advisor (Dr. John Kelly, University of Kansas) and I set up a large (~40,000 plant) experimental evolution study, using a primarily outcrossing North American wildflower, Mimulus guttatus (monkeyflower). These cute yellow-flowered plants possess highly variable morphology and life history traits and are primarily pollinated by native bumblebees.
As a starting point for experimental evolution, we used seed from a large source population to create two experimental treatment groups, each containing two replicate populations: “Bee” populations (B1, B2) received abundant bumblebee (Bombus impatiens) pollinators while “No Bee” populations (A1, A2) experienced a complete lack of pollinators. We allowed these four initially equivalent populations to evolve for 5 generations under the two pollination treatments in a greenhouse.
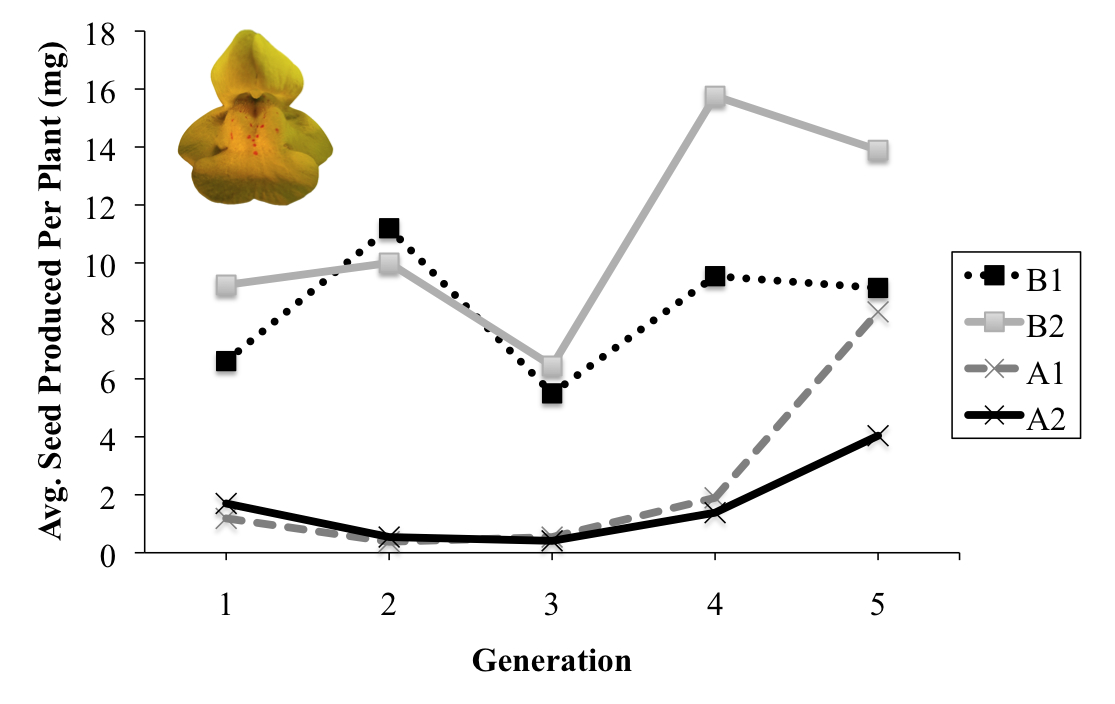
Population fitness over 5 generations of evolution. B1 and B2 are Bee populations, A1 and A2 are No Bee populations. Inset – M. guttatus flower.
We recorded plant phenotypes and genotypes in the final generations of the study and were impressed with the speed of the evolution observed within and among the treatments. Populations without pollinators (No Bee) displayed low and declining fitness (measured by seed production) in the early generations of the study but then rebounded, culminating in a ten-fold increase in self-fertilization capacity compared to the initial ancestral population (see figure at right). This pattern—a rapid increase of adapted genotypes after an environmentally induced fitness crash—is consistent with the theory of evolutionary rescue. Thus, at least under our experimental conditions, we demonstrated that rapid mating system evolution via increased selfing capacity is a potential adaptive strategy for plant populations experiencing pollinator loss. Genotyping of our experimental plants at multiple loci revealed rapid allele frequency changes occurring in the No Bee populations, which were correlated with the evolving phenotypes.
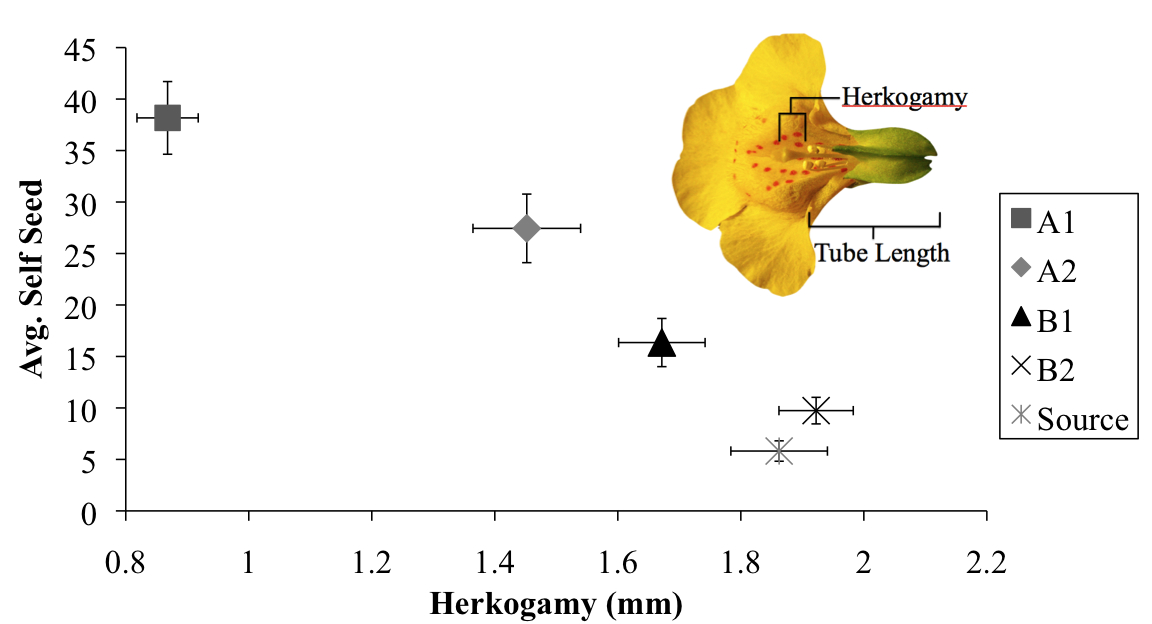
Bivariate plot of the negative relationship between mean herkogamy and mean self seed by experimental population after 5 generations. Error bars are +/- 1 SEM. B1 and B2 are Bee populations, A1 and A2 are No Bee populations, and Source is the original population without treatment. Inset—cross-section of flower, with floral measurements.
Additionally, we identified a potential mechanism that may have contributed to the observed increases in selfing in the No Bee populations. Across populations, we found that selfing ability was correlated with herkogamy, the measured distance separating the male and female reproductive parts of a flower. Populations that had increased selfing rates had reduced herkogamy (see figure at left).
Our experimental evolution study demonstrates that rapid adaptation of reproductive strategy in response to pollinator loss is possible, but we also know that it comes with a cost. Selfing is complete inbreeding – and one disadvantage of inbreeding is a loss of genetic diversity, making populations vulnerable to extinction if further environmental perturbations occur.
Future directions
Current work on the monkeyflower mating system evolution project focuses on identifying specific regions of the genome that
evolve during a mating system transition from outcrossing to selfing. To accomplish this, we are developing a novel association technique, similar to standard marker-based QTL mapping, that targets highly divergent genomic regions from full-genome sequencing of experimental populations. Forthcoming results will help us understand how plant genomes respond to selection for increased selfing.
I originally became interested in mating system evolution during an NSF REU experience as an undergraduate, learning about northern cardinal (Cardinalis cardinalis) mate guarding and extra-pair paternity behavior. At MSU, I’m going back to my avian roots by developing a collaborative project with Dr. Tom Getty, graduate student Cara Krieg, and Dr. Lindsay Walters (Northern Kentucky University) to look at plastic responses of mating systems in the house wren (Troglodytes aedon).
For further information about these and other research projects, contact Sarah at bodbyl at msu dot edu or visit her website. Sarah also manages the NSF KBS GK-12 Bioenergy Sustainability project, a graduate training and research outreach program; check out their website here.
