
This week’s BEACON Researchers at Work blog post is by University of Washington graduate student Peter Conlin.
Natural selection produces an organism whose phenotype is well matched to its environment. Under a constant environment there should be a single optimum, but what happens in a varying environment when different phenotypes are favored at different times? Environmental change can be seen as a challenge because it may disrupt the match between phenotype and environment, leading to a decrease in fitness. One solution to this mismatch problem is for individual organisms to condition their phenotype on the state of their environment.
The ability of a single genotype to alter its phenotype in response to changes in the environment is called phenotypic plasticity. Phenotypic plasticity can play many important roles in evolution – it can increase fitness, generate novelty, and plays a role in structuring ecological communities. This concept was first applied to changes in morphology but has more recently been extended to include a diversity of environmentally induced changes.
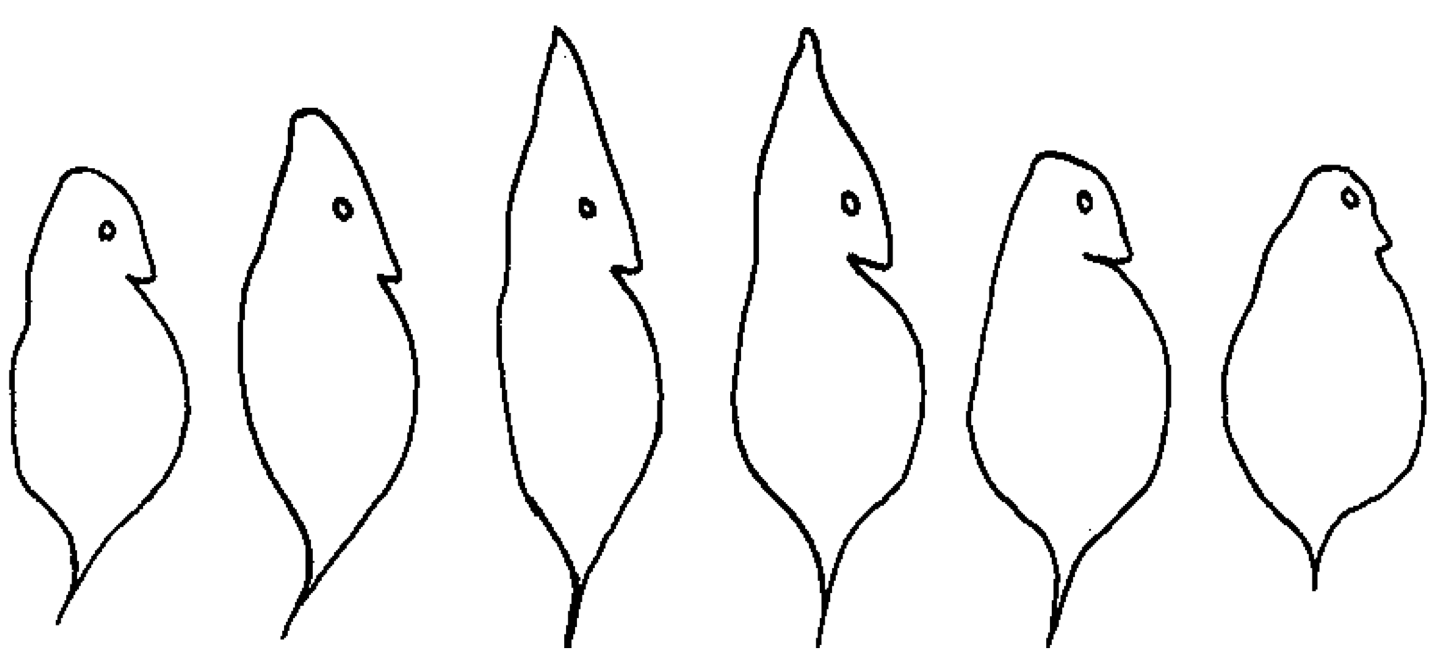
The development of a defensive head shield and tail spines in Daphnia. The first described and arguably cutest example of phenotypic plasticity (from Woltereck, 1909).
A classic example of adaptive phenotypic plasticity, and the first described, comes from the water flea, Daphnia. When grown in the presence of a predator Daphnia will grow defensive head shields and tail spines that are thought to increase their chance of survival by deterring predators.
Phenotypic plasticity is found across all levels of biological complexity and theory predicts that plasticity can be adaptive when the following criteria hold:
Organisms experience different environments (spatially or temporally).
- Different environments favor different phenotypes.
- The environmental cue provides reliable information about selective conditions.
The third criterion is especially important because in some cases, the phenotypic change must precede the selective conditions. In the case of Daphnia, the defensive head shield must grow before encountering a predator (if it is to be effective) and growing a head shield in the absence of a predator is thought to be costly. Daphnia are known to use chemicals released by their predators as a cue to grow a head shield. Interestingly, researchers have found that an increase in water temperature will elicit the same defensive response in some species!
A vast literature on phenotypic plasticity exists from studies of natural and laboratory populations, much of it from just the past 20-30 years. Experiments have demonstrated that plasticity is an evolvable trait, that plasticity can be directly selected, and have given us a better understanding of the complex genetics of phenotypic plasticity. Even so, a great number of theoretical predictions about the evolution of plasticity remain untested. My research in Ben Kerr’s lab uses an experimental evolution approach to understand the role of environmental cue reliability in the evolution of phenotypic plasticity and what genetic changes occur as plasticity evolves.

The phenotypic plasticity team. From left to right: Joseph Marcus, Samuel Reed, and Peter Conlin.
We are working with a strain of the budding yeast, Saccharomyces cerevisiae, from Will Ratcliff and Mike Travisano at the University of Minnesota that forms multicellular clusters, a phenotype caused by incomplete cell separation. The phenotype we are focusing on is the size of the yeast cluster. We chose this because cluster size varies across isolates and clusters can be easily size-sorted by centrifugation due to differential rates of sedimentation.
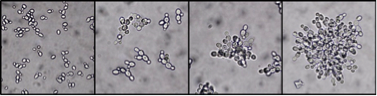
Multicellular yeast clusters. This phenotype is found in many natural isolates but also readily evolves when yeast are grown under selection for rapid settling. Multicellular yeast clusters have also recently been evolved by Andrew Murray’s lab under selection for improved utilization of a public good.
In the experiment yeast clusters are grown in a pair of alternating cue environments referred to as E1 and E2. We can favor the evolution of plasticity by pairing the different environments with opposing selective conditions (propagating either the top or bottom of the culture after centrifugation). Cue reliability is high when E1 and E2 are uniquely paired to top or bottom selection, respectively. We can test theoretical predictions about the importance of cue reliability by deviating from the 1:1 correlation between environment and selection. Our hypothesis is that phenotypic plasticity is more likely to evolve when cue reliability is high. We hope that this work will shed light on the conditions that favor the evolution of phenotypic plasticity and the underlying mechanisms for achieving plasticity.
For more information about Peter’s work, you can contact him at pconlin2 at u dot washington dot edu.
