
This week’s BEACON Researchers at Work blog post is by University of Idaho postdoc Wesley Loftie-Eaton.
 With all the thanks to BEACON for funding and Dr. Eva M. Top for her support and guidance, my latest foray into the world of plasmids explores a question that I have been thinking about for nearly a decade: How, and under what conditions does plasmid host range evolve when the plasmids do not encode any genes obviously beneficial to the host or when there is no strong external selection for their maintenance?
With all the thanks to BEACON for funding and Dr. Eva M. Top for her support and guidance, my latest foray into the world of plasmids explores a question that I have been thinking about for nearly a decade: How, and under what conditions does plasmid host range evolve when the plasmids do not encode any genes obviously beneficial to the host or when there is no strong external selection for their maintenance?
For the non-(micro)biologists reading this, plasmids are self-replicating DNA molecules that occur in bacteria and are quite often capable of spreading horizontally between diverse bacterial genera, making them central to horizontal gene transfer and rapid bacterial adaptation. With that said, we generally tend to study plasmids in context of their role in spreading antibiotic resistance genes and virulence factors between pathogenic bacteria. The focus on this phenomenon is driven by the immense public health threat brought about by antibiotic resistance (Kahrstrom, 2013; Obama, 2014). As a result, most of our efforts to understand the evolution of plasmid persistence and consequently host range include (periodic) conditions that render the plasmids beneficial to their hosts, such as the presence of antibiotics to which the plasmids encode resistance (Bouma and Lenski 1988; Modi et al. 1991; Modi and Adams 1991; Dahlberg and Chao 2003; Dionisio et al. 2005; Heuer et al. 2007; De Gelder et al. 2008; Sota et. 2010; San Milan et al 2014a; San Milan et al. 2014b; Harrison et al., 2015). While there is no arguing the contribution of plasmids towards the spread of antibiotic resistance genes and virulence factors, we have to remember that many plasmids in nature lack genes that encode host-beneficial functions (Brown et al. 2013, Loftie-Eaton and Rawlings, 2012). Because these so-called ‘cryptic’ plasmids lack an obvious benefit to their host there is no strong external selection for their maintenance and thus it is unclear what conditions can facilitate evolution of their host range.
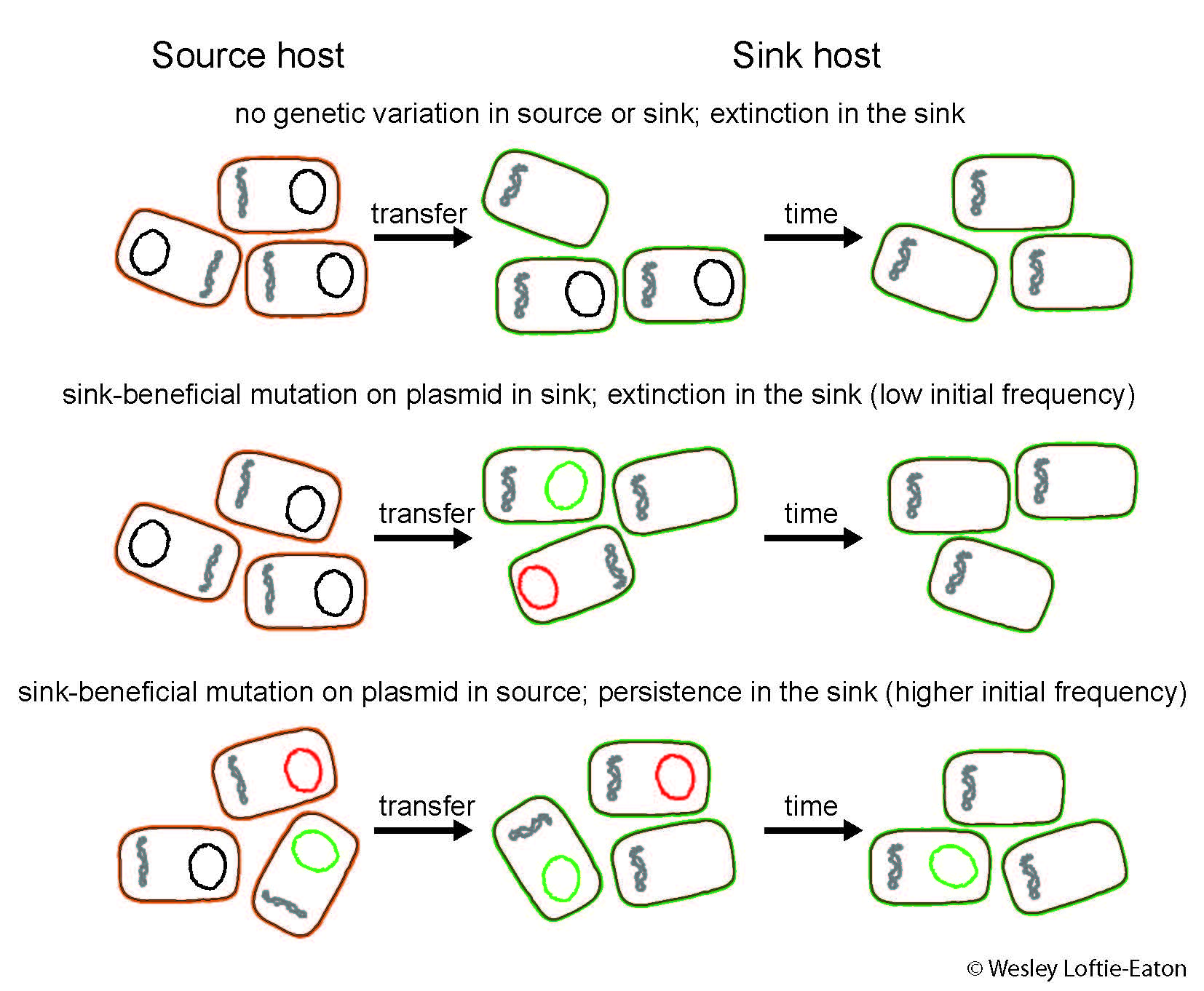
Fig. 1 An illustrated hypothesis of how genetic variation in the genome of a plasmid from a source host can facilitate greater persistence in a sink host. Plasmids with mutations that lead to increased persistence in the sink hosts are indicated in green, whereas plasmids with mutations that are deleterious/neutral in the sink population are indicated in red.
Borrowing from ecological source-sink theory my colleagues and I have formulated a hypothesis that may help us understand ecological conditions that facilitate plasmid host range evolution in the absence of strong external selection. In source-sink theory a source is a habitat in which the net birth rate within a population is positive, whereas a sink is a habitat in which the net birth rate is negative and population persistence relies on immigration from a source (Pulliam, 1988). Evolutionary source-sink theory predicts that adaptation to a new habitat is unlikely to occur if the reproductive rate in that environment is low. However, given sufficient genetic variation from a source population, mutant alleles may increase reproductive success in the sink habitat following immigration (Dennehy et al, 2010). The “habitat” of a plasmid is its bacterial host. A source host of a plasmid is thus a bacterial species in which the plasmid is capable of long-term persistence, whereas a sink host is a bacterial species from which the plasmid is rapidly lost due to high segregational loss, reduced host growth rates brought about by high plasmid cost, or inefficient horizontal spread. However, from previous research we know that as little as one mutation in the plasmid can result in increased persistence (Sota et al, 2010; Loftie-Eaton et al, submitted). Thus we hypothesize that genetic variation in the genome of a plasmid from a source host can facilitate greater persistence in a sink host (Fig. 1).

Fig. 2 Source-sink population dynamics to study plasmid host range evolution can be achieved and controlled in chemostats. This is done by creating an environment that precludes the source host from growing while allowing for continuous growth of the sink host. In such an environment the only way for the plasmid to propagate is to transfer to the sink host before the source host is diluted out. In theory, plasmids with sink-beneficial mutations that conjugated from the source to the sink will be able to persist for a longer period of time than the ancestral plasmid or plasmids with sink neutral/deleterious mutations. Plasmids with different alleles are indicated in different colours.
To test our hypothesis we put together a team with experience in microbiology, ecology, evolutionary theory and mathematical modeling and devised an experimental design that is driven by explicit theoretical predictions to be tested using empirical and computational methods. The plasmid we are using is a cryptic conjugative plasmid that was isolated from the pathogenic bacterium Bordetella pertussis (causes whooping cough; Kamachi et al., 2006). The source host that we are using is an E. coli that is quite capable of maintaining the plasmid in the absence of strong selection, whereas the sink host, Shewanella oneidensis, is not (Yano et al., 2013). The source host, however, is an auxotrophic mutant whose growth can be selectively inhibited by excluding a nutrient essential to it, but not to the sink host. Thus, in a constantly diluting environment such as a chemostat in which the source host cannot grow, the only way for the plasmid to persist is to conjugate to and propagate in the sink host (Fig. 2). However, since the plasmid is maladapted to the sink host it will not be able to persist for long unless, in accordance with our hypothesis, there are mutant alleles in the genome of the plasmid that allows for increased reproductive success in the sink host. Based on current estimates of the parameters describing persistence of the plasmid in a monoclonal population of the sink host, our preliminary modeling predicts that mutant alleles in the plasmid genome that b
ring about a 10% decrease in plasmid cost or a 100-fold decrease in segregational loss rate can, in either case, result in the plasmid persisting 500 generations more than the ancestral plasmid in the sink host, thus supporting our hypothesis in silico.
For me, as a plasmid biologist dabbling in evolution, this project is super exciting. We will for the first time provide experimentally derived insight into the evolution of host range of cryptic plasmids. But moreover, having a tractable source-sink experimental system combined with computational methods will open avenues to empirically test different theoretical source-sink migration dynamics. For example, our results will guide future studies with other bacteria and plasmids and diverse experimental conditions, including growth in biofilms, to explore how source-sink and other population dynamics can explain some of the observed abundance of plasmids in natural bacterial populations. This is important because to successfully control the spread of antibiotic resistance, which is rapidly becoming a global health crisis, we need to understand the persistence, spread and evolution of all plasmids, not just resistance plasmids.
Acknowledgements
This work would not be possible without my excellent team members which includes Dr. Eva Top and Genevieve Metzger at the University of Idaho, and Dr. John Mittler and Jacob Bayless da Costa at the University of Washington, nor would it be possible without funding from BEACON.
References
- Bouma J. E., R. E. Lenski. Evolution of a bacteria/plasmid association. Nature 335:351-352.
- Brown, C. J., D. Sen, H. Yano, M. L. Bauer, L. M. Rogers, G. A. Van der Auwera, and E. M. Top. 2013. Diverse broad-host-range plasmids from freshwater carry few accessory genes. Appl Environ Microbiol 79:7684-7695.
- Dahlberg C., L. Chao. Amelioration of the cost of conjugative plasmid carriage in Eschericha coli K12. Genetics 165:1641-1649.
- De Gelder L., J. J. Williams, J.M. Ponciano, M. Sota, E. M. Top. Adaptive plasmid evolution results in host-range expansion of a broad-host-range plasmid. Genetics 178:2179-2190.
- Dennehy, J. J., N. A. Friedenberg, R. C. McBride, R. D. Holt, and P. E. Turner. Experimental evidence that source genetic variation drives pathogen emergence. Proc Biol Sci 277:3113-3121.
- Dionisio F., I. C. Conceição, A. C. Marques, L. Fernandes, I. Gordo I. The evolution of a conjugative plasmid and its ability to increase bacterial fitness. Biol Lett 1:250-252.
- Harrison E., D. Guymer, A. J. Spiers, S. Paterson, M. A. Brockhurst. Parallel compensatory evolution stabilizes plasmids across the parasitism-mutualism continuum. Curr Biol 25(15):2034-2039.
- Heuer H., R.E. Fox, E.M. Top. Frequent conjugative transfer accelerates adaptation of a broad-host-range plasmid to an unfavorable Pseudomonas putida host. FEMS Microbiol Ecol 59:738-748.
- Kahrstrom, C. T. 2013. Entering a post-antibiotic era? Nat Rev Microbiol 11:146.
- Kamachi, K., M. Sota, Y. Tamai, N. Nagata, T. Konda, T. Inoue, E. M. Top, and Y. Arakawa. 2006. Plasmid pBP136 from Bordetella pertussis represents an ancestral form of IncP-1beta plasmids without accessory mobile elements. Microbiology 152:3477-3484.
- Loftie-Eaton, W. and D.E. Rawlings. 2012. Diversity, biology and evolution of IncQ-family plasmids. Plasmid 67:15-34.
- Loftie-Eaton W., H. Yano, S. Burleigh, R. S. Simmons, J. M. Hughes, L. M. Rogers, S. S. Hunter, M. L. Settles, L. J. Forney, J. M. Ponciano, E. M. Top. Evolutionary paths that expand plasmid host-range: implications for spread of antibiotic resistance.
- Modi, R. I., J. Adams. 1991. Coevolution in bacterial-plasmid populations. Evolution 45:656-667.
- Modi R. I., C. M. Wilke, R.F. Rosenzweig, J. Adams. Plasmid macro-evolution: selection of deletions during adaptation in a nutrient-limited environment. Genetica 84:195-202.
- Obama, B., The White House, Office of the Press Secretary. 2014. Remarks by the president in state of union address Washington, D.C.: Retrieved from http://www.whitehouse.gov/the-press-office/2014/01/28/president-barack-obamas-state-union-address.
- Pulliam, H.R. Sources, sinks, and population regulation. Amer Nat 132:652-661.
- San Millan A., K. Heilbron, R. C. Maclean. Positive epistasis between co-infecting plasmids promotes plasmid survival in bacterial populations. ISME J. 8:601-612.
- San Millan A., R. Peña-Miller, M. Toll-Riera, Z.V. Halbert, A. R. McLean, B.S. Cooper, R. C. MacLean. Positive selection and compensatory adaptation interact to stabilize non-transmissible plasmids. Nat Commun. 5:5208.
- Sota, M., H. Yano, H. M, Julie, G. W. Daughdrill, Z. Abdo, L. J. Forney, and E. M. Top. Shifts in the host range of a promiscuous plasmid through parallel evolution of its replication initiation protein. ISME J 4:1568-1580.
- Yano, H., L. M. Rogers, M. G. Knox, H. Heuer, K. Smalla, C. J. Brown, and E. M. Top. Host range diversification within the IncP-1 plasmid group. Microbiology 159:2303-2315.
For more information about Wesley’s work, you can contact him at wesleyl at uidaho dot edu.
