
This week’s BEACON Researchers at Work blog post is by MSU graduate student Samuel Perez.
 The study of morphological traits, physical features that are directly observable and measurable, is important for the study of evolution, and play a central role in Darwin’s theory of natural selection. Organisms interact with their environment using their morphological traits to do tasks like obtain energy from limited resources, avoid predation, maintain homeostasis, and reproduce. The traits involved in these tasks often have differences between individuals (variation) in a population, and some traits have more variation than others. If those morphological differences lead to differences among individuals in the number of offspring, then natural selection results and evolution at the population level can occur; this process is known as adaptation.
The study of morphological traits, physical features that are directly observable and measurable, is important for the study of evolution, and play a central role in Darwin’s theory of natural selection. Organisms interact with their environment using their morphological traits to do tasks like obtain energy from limited resources, avoid predation, maintain homeostasis, and reproduce. The traits involved in these tasks often have differences between individuals (variation) in a population, and some traits have more variation than others. If those morphological differences lead to differences among individuals in the number of offspring, then natural selection results and evolution at the population level can occur; this process is known as adaptation.
However, if a formerly-adaptive trait no longer contributes to the fitness of a population, perhaps due to changing environmental conditions, then what happens to that trait? In that case, there can be two possibilities: either the trait will become reduced and lost as it accumulates mutations and resources are invested in other, more essential features that can contribute to adaptation, or the trait can be modified towards a new function. A striking example of trait loss in animals is the loss of flight, which has occurred in two major groups of animals that have developed flight (insects and birds). Some insects have lost their wings altogether, while penguin wings have been co-opted for swimming.

A.thaliana flower. Note that pollen from the mature short stamen does not reach the pistil (center) part of the flower.
Pic: Jürgen Berger/Max Planck Institute for Developmental Biology, Tübingen, Germany
I’m interested in knowing how much variation is observed in a trait undergoing loss at the population level, the genetic basis of trait loss, why nonfunctional traits are sometimes maintained, and how formerly adaptive traits can be shifted towards new functions (co-option). I study trait loss in the model plant Arabidopsis thaliana (thale cress), of the Brassicaceae family, which has almost 4,000 species and includes species like mustard, cabbage, canola, and radishes. A conserved feature in Brassicaceae is that flowers have four long stamens (male reproductive parts) and two short stamens.
However, A. thaliana produces more than 98% of its seeds through self-pollination, and the short stamen anthers, where the pollen is produced, never come near the stigma, where the pollen needs to be deposited to fertilize the seeds (see picture above). Previous experiments by Anne Royer in which short stamens and long stamens were removed showed that the short stamens do not contribute significantly to self seed production, making it an example of a nonfunctional trait. Anne also showed that the short stamens have begun to be lost in some populations of A. thaliana, and this loss exhibits a latitudinal cline in the native European range: stamen loss is common in the Mediterranean but rare in Scandinavia.

A. thaliana flowers with six stamens (left), five stamens (center), and four stamens (right).
Many animal and plant traits that exhibit a latitudinal cline also show an altitudinal cline as well; lower latitudes correspond to low altitudes, and higher latitudes correspond to lower altitudes due to similarities in climate. To see if this is true for stamen loss in A. thaliana, I counted short stamens in 15 natural populations from N.E. Spain that ranged from 109-1688 m in altitude. I found that, in parallel to the latitudinal cline, populations at lower altitudes had more short stamen loss than populations at higher altitudes (see graph below). These parallel latitudinal and altitudinal clines suggest that there is a common source of natural selection at work, either directly on short stamens or through a correlated trait.
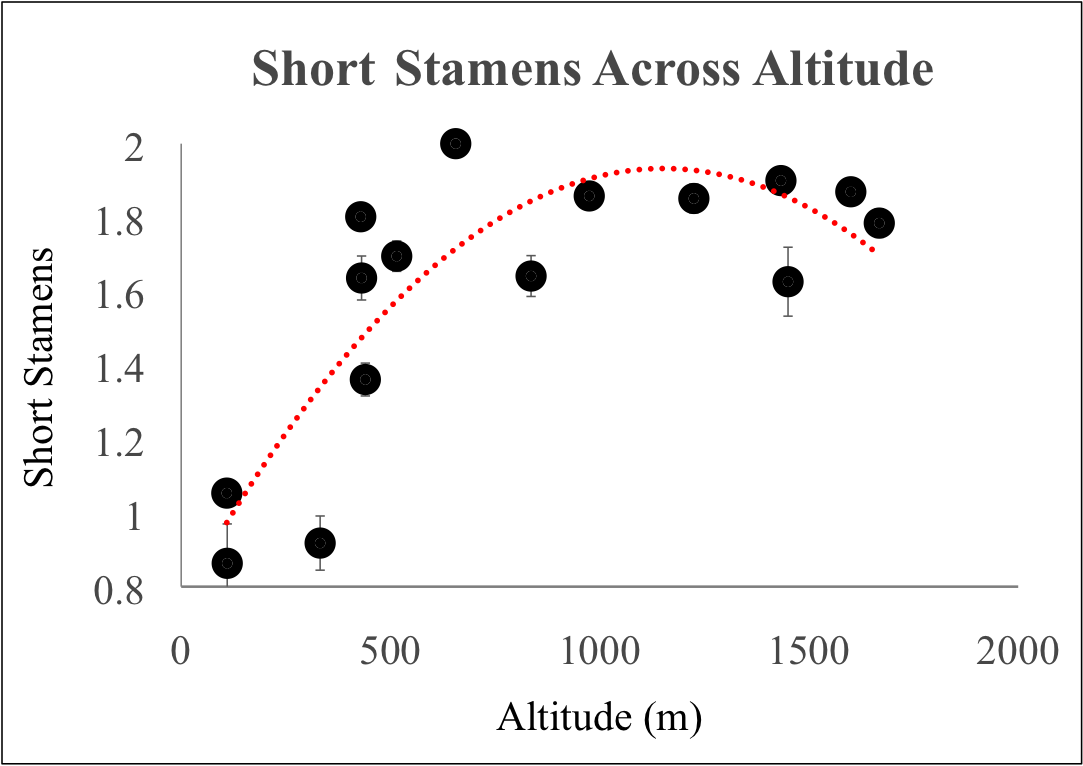
Mean short stamen number (±1 SEM) from 15 populations in N.E. Spain. Quadratic fit is provided in red.

One of the advantages of studying lines from multiple populations of Arabidopsis thaliana is more potential for phenotypic and genetic variation across floral traits. Green circle diameter = 1.9 cm.
I also measured stamen loss from a series of inbred line crosses between two populations from Sweden and two populations from Spain to look at the genetic basis of stamen loss across the whole genome. We found that there was genetic variation in stamen loss within one of the Italian populations. The F1 generation showed evidence of recessive genes underlying stamen loss, the F2 and backcrosses suggested that there were epistatic interactions between the genes responsible for stamen loss. The gene interactions may be slowing the evolution of stamen loss even when genes have additive effects.
My thesis research will focus on furthering the understanding of the selective and genetic mechanisms underlying stamen loss. I will examine the fitness effects of stamen loss, both direct effects through costs of production of short stamens, as well as correlated traits that may be involved with adaptation to high altitude and latitude. I will conduct genome-wide association mapping studies (GWAS) using the whole-genome sequence data to look for the genes responsible for the latitudinal cline in stamen loss.
For more information about Sam’s work, you can contact him at perezsa4 at msu dot edu.
