
This week’s BEACON Researchers at Work blog post is by University of Texas at Austin graduate student Rayna Harris.
Research in Hans Hofmann’s lab at UT Austin is best known for its studies of the neurogenomic basis of male social behavior, but Hans and his entourage have never shied from using comparative approaches to study the evolution of social behavior (listen to Hans on NPR’s Academic Minute). My research in the Hofmann lab has fallen into the second category, as two of my favorite comparative experiments investigated 1) the behavioral evolution from a monogamous to polygynous mating system and 2) the molecular evolution of the neuropeptide pro-opiomelanocortin (pomc).
Mating System Evolution in Herichthys Cichlid Fishes

A H. cyanoguttatus pair defends their territory.
In 2010 and 2011, I investigated the neuroendocrine mechanisms that underlie differences in social behavior between two closely related North American cichlid fishes (1). Herichthys cyanoguttatus, the Rio Grande cichlid, lives in rivers and drainages of the Gulf Coast of northern Mexico and southern Texas. They are sexually monomorphic (see photo at right), monogamous, and provide bi-parental care for offspring, often over multiple breeding cycles. Herichthys minckleyi, the Cuatro Ciénegas cichlid, lives in the pristine spring-fed ponds and streams in the desert valley of Cuatro Ciénegas in northern Mexico. They are sexually dimorphic and polygynous, with males providing little paternal care.
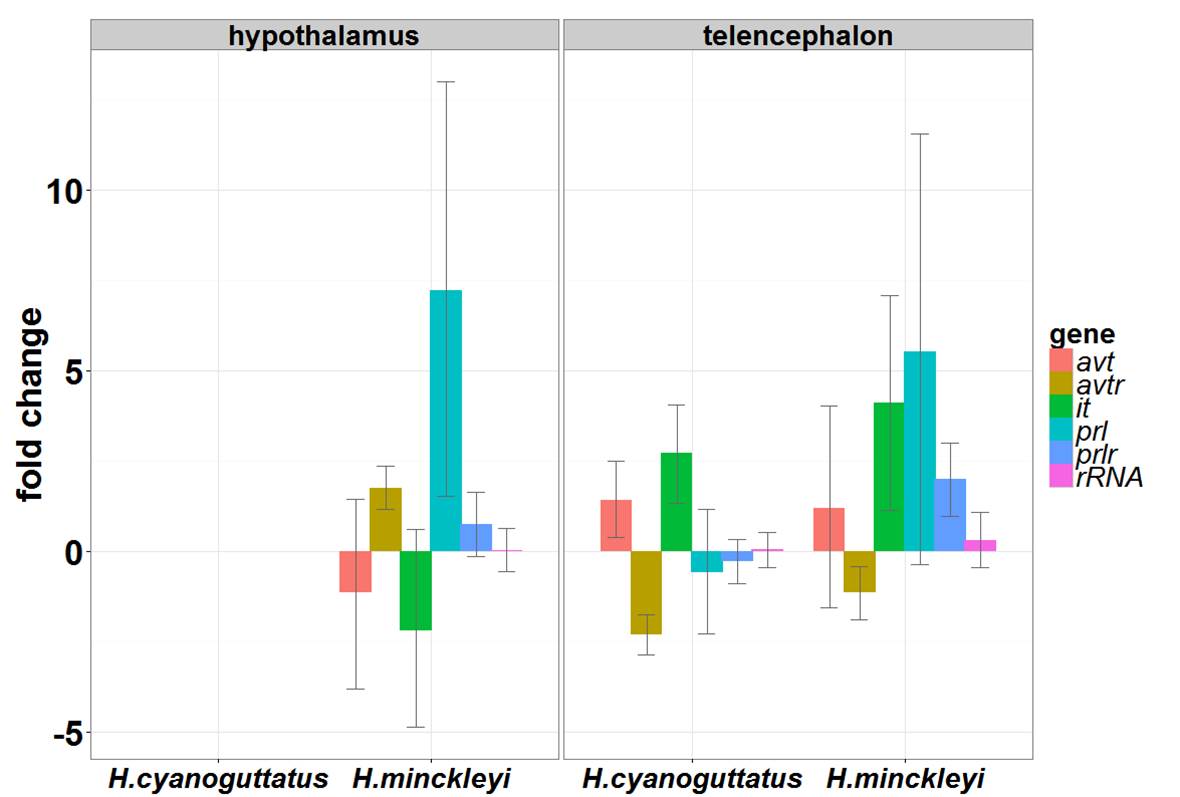
Neuroendocrine changes. This graph shows how expression of arginine vasotocin (avt) and its receptor (avtr), isotocin (it), prolactin (prl) and its receptor (prlr) differ by brain region and species as compared to hypothalamic expression in H. cyanoguttatus. 18S rRNA expression is stable. Bars show median expression (log2fold change) and whisker are 95% confidence intervals.
We collected blood, brains and gonads in the “field” (our field stations were set up on the running path along Shoal Creek Trail just north of 6th Street in downtown Austin and in two outdoor pools at UT’s J.J. Pickle Research Campus). In the lab we micro-dissected the brains and processed the blood for quantitative real-time PCR and hormone assays, respectively. We found that two circulating androgens are differentially regulated between males of each species. I analyzed my qPCR data with an awesome new R package called MCMC.qpcr and created bar graphs to visualize the species- and brain region- differences relative to H. cyanoguttatus hypothalamic expression (see figure at left). A few significant results include increased expression of prolactin (prl) in the hypothalamus and prolactin receptor (prlr) in the telencephalon H. minckleyi, indicating increased activity of the prolactin pathway in this species. Next, I used a correlation network approach to identify patterns of conserved endocrine activity in these two species (below). This covariance network suggests a relationship between circulating androgen levels and neuroendocrine receptor expression in the brain. Up-regulation of gene networks, not individual genes, appears to be important for driving changes in behavior. Future research is needed to disentangle neural differences associated with habitat independent of social behavior.

Covariance network. This network shows the relationship between the androgens testosterone and 11-ketotestosterone (11-KT) and gene expression in the telencephalon (TEL) and hypothalamus (HYP) across all individuals in the analysis.
This study focused on prolactin, vasopressin, and isotocin pathways, but other research in our lab was investigating the role of the melanocortin system in regulating social behavior and other phenotypic traits in cichlid fishes.
Molecular Evolution of the Pro-opiomelanocortin Gene Family
In 2011, the genomes of five African Cichlid species were made publicly available, and I became engrossed with understanding to which extent the melanocortin system underlies polymorphisms in color, behavior, and physiology across species. I designed a research project to examine the molecular evolution the neuropeptide pro-opiomelanocortin (pomc) gene family (2). Most teleosts have a duplicate pomc gene from the teleost whole genome duplication (pomc β), but the pomc α paralog recently duplicated in multiple lineages independently (pomc α1 and α2). I was isolating and sequencing these paralogs in A. burtoni when I obtained a curious result in the pomc β gene: a portion of the α-melanocyte stimulating hormone (α-MSH) region was duplicated! After confirming that it wasn’t a technical artifact (the duplication is present in African- and American cichlids and damselfish), I devoured the literature and learned that the α-MSH region of pomc gene had four other times in evolution. I had just uncovered the fifth example, so I named the region “ε-MSH” to follow the established nomenclature on naming peptides with a melanocortin binding site (cleaved peptide sequence: S Y R M E H F R W G K P A G L K M R E P K L K A R S D E). Next, I used bioinformatic approaches to understand how changes in regulatory sequence gave rise to differential tissue expression of pomc orthologs and paralogs. This analysis of subfunctionalization of duplicated genes inspired me to use functional approaches to study genome regulation as part of my dissertation, which aims to examine evolution of steroid hormone response elements in the genome. So
, stay tuned for more on that the subject of cis-regulatory evolution of steroid hormone receptors.
Outreach & Networking

International networking. Hans and I (center) had a great time participating at the conference and workshop in Bergen, Norway.
I gained a lot of practical experience working on the two projects above, but perhaps even more importantly, they provided many opportunities for honing my writing and speaking skills. I presented the mating system evolution story to my peers at the 2nd Annual Brain, Behavior & Evolution Annual Symposium and to the public during a Darwin Day outreach event and on the radio program “They Blinded Me with Science”. I presented the molecular evolution story at a conference in Bergen, Norway and at the Gordon Research Conference: Genes & Behavior. All of these activities provided a stimulating environment to discuss my research with the intellectual and the curious, and they opened new doors for opportunities in research, teaching, and outreach. Who knows, maybe one day I’ll give a TED talk about the role of molecular evolution in shaping behavioral polymorphisms.
- Oldfield RG, Harris RM, Hendrickson DA, Hofmann HA (2013) Arginine Vasotocin and Androgen Pathways are Associated with Mating System Variation in North American Cichlid Fishes. Hormones and Behavior 64: 44–52.
- Harris RM, Dijkstra PD, Hofmann HA (2014) Complex structural and regulatory evolution of the pro-opiomelanocortin gene family. General and Comparative Endocrinology 195,: 107–115
For more information about Rayna’s work, you can contact her at rayna dot harris at utexas dot edu or like the Hofmann lab Facebook page for research updates.
