
Cross-posted from UT postdoc Art Covert’s blog, Covert Science(ish)
Names are generally very arbitrary things. In the words of The Bard: “A rose by any other name would smell as sweet.” This may be true for roses, but for mutations, names carry a great deal of meaning. Beneficial mutations are good, they improve an organism’s chances for survival (i.e. raise its fitness). Deleterious mutations are bad, they reduce an organisms chances for survival (lower fitness). One type of mutation is distinctly sweet, the other distinctly wretched.
Mutations are assigned their names when they first appear in a population. A mutation that improves fitness is assumed to be good and a mutation that lowers fitness are assumed to be bad. These name assignments seem nice and neat and simple, but they are predicated on the assumption that a mutation’s fate is unalterable. Good mutations are kept, bad mutations are purged by selection, very neat and tidy just as we scientists prefer it.
The picture becomes murkier when you allow for a second mutation that appears on the same genome as the first. That second mutation has the potential to alter the effect of the first. Good mutations may become bad and bad mutations become good (Figure 1).
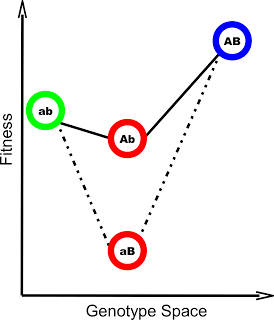
Figure 1: An example of the two mutation system described above (From Covert et al 2013). Both mutations A and B individually are deleterious, but A and B together are extremely beneficial.
So if a second mutation can alter the effect of the first then all bets are off, right? Classical evolutionary theory doesn’t think so, and has held that such infrequent occurrences are probably unimportant. Mutations are rare; getting two mutations is even rarer. Getting the right two mutations, close together, to not only ameliorate a deleterious mutation, but to actually open up new beneficial mutations that weren’t accessible before? Pishaw! You get better odds at the blackjack table! So the literature generally holds that bad mutations stay bad, and good mutations stay good, and that’s how it’s been in evolutionary theory for a long time.
Evolution isn’t about playing one hand of blackjack though, its about playing lots and lots of hands, over a very long period of time. On a long enough timescale, you will eventually see one of these rare chance events. Deleterious mutations are extremely common, a genotype with a deleterious mutation will persist in the population for a few generations before it’s purged. In that short time, if one of that genotype’s progeny acquires a second mutation their is a small chance that the two mutations together will actually be good. There is also a possibility that in addition to ameliorating the deleterious effect, the pair of mutations also pushes the population’s genotypes closer to other beneficial mutations. These super-compensatory mutations are rare, but have a huge impact when they finally emerge.
The question is not “if” or “when”, but of how important these super-compensatory mutations are. Until now this question has been open, but now my colleagues and I have begun to shed light on it. Using self-replicating computer programs (digital organisms) we examine the effects of deleterious mutations in two ways that are not possible with organic study systems.
In our first set of experiments, we replace all deleterious mutations with beneficial or neutral mutations making it impossible for deleterious mutations to appear at all. In contrast, mutations in the population normally occur completely at random with the vast majority of them having deleterious or lethal fitness effects. Surprisingly, when we compare the no-deleterious populations to normal populations we find that the normal populations have much much higher fitness. So, at least some immediately deleterious mutations in the long run are creating a net benefit. But is it all deleterious mutations that are important or just a few?
With our second set of experiments we undid every instance of a deleterious mutation on the lineage from the starting organism to the most successful genotype in each population. Undoing these mutations asks the question “what would have happened without this one mutation?” Throughout all of the populations with deleterious mutations we found rare instances of super-compensatory deleterious mutations, which also lead populations to previously inaccessible beneficial mutations. In other words, occasional steps that are not immediately beneficial may lead to huge rebounds.
This isolation of historically significant super-compensatory mutations is what sets our work apart from other works on the role of deleterious mutations. We actually identify which deleterious mutations are leading to higher fitness in the long run and we measure their impact on the long-term evolution of the population.
The most startling thing we found when we looked at individual super-compensatory mutations, is that they occur in minuscule quantities. Out of 50 replicate populations, each experiencing 45,000 generations of evolution, we found only 36 super-compensatory mutations. Of those 36, only 11 were found to be necessary for the populations’ continued evolution. But theses 11 super-compensatory mutations fundamentally altered the outcome of evolution. Without these extremely rare events the evolutionary process stalls out, possibly for very long periods of time.
So the story is not told by the name alone. Every once in a while, a deleterious mutation will interact with other mutations in a way that makes them more beneficial together; interactions such as these are known in the literature as “epistatic” mutations. These rare, but historic events are pathways to essential variation in evolving populations that may become evolutionarily stuck otherwise. Furthermore, it’s impossible to predict *which* deleterious mutation will be the lucky one. But one thing from my work is clear: deleterious mutations do provide essential variation needed by evolving populations.
Covert, Lenski, Wilke and Ofria (2013) Experiments on the role of deleterious mutations as stepping stones in adaptive evolution. PNAS doi: 10.1073/pnas.1313424110
